
Our Research
We investigate the fundamental organisational principles and evolutionary dynamics of the nuclear compartment across eukaryotes using comparative genomics, cell biology and experimental evolution.
Read about some of our ongoing projects below!
We are generally interested in how evolution shapes cell biology. As part of our broader efforts, we are developing screens and reporter tools to be used across evolutionarily close and distant organisms such as S. pombe, the budding yeast Saccharomyces cerevisiae, and the slime mould Physarum polycephalum.
An important component of our toolkit is ultrastructure expansion microscopy (U-ExM). We have already optimised the method for its use in the model yeasts S. pombe and S. cerevisiae (together with the Centriole Lab and Loewith Lab) and are aiming to adjust the protocol to work on complex samples containing multiple species.
Understanding Opisthokont nuclear organization using orthology-based reconstruction of regulatory networks



The nucleus is universally present across eukaryotes but exhibits a great variation in how it is structurally organised. Despite significant advances in genome sequencing, computational tools, and microscopy, little is known about the principles underlying the evolutionary history of nuclear structural organisation that can explain its observed extant diversity. What is known through studies in tractable model systems is difficult to generalise given the limited phylogenetic and phenotypic coverage.
We propose to bridge this gap in knowledge by setting up a workflow to infer protein orthologs using Hidden Markov Models, identify protein interaction modules, and create genotype-to-phenotype maps in uncharacterised Opisthokonts based on data from model systems.
This will also be a valuable resource for evolutionary cell biologists.
This project is a collaboration with Yannick Schwab (EMBL), Omaya Dudin (EPFL), and Tom Williams (University of Bristol)
In the face of different selection pressures, some eukaryotes have evolved to replicate genetic material without generating new daughter cells, resulting in multinucleate cells. The dynamics and synchronicity of nuclear divisions in multinucleate cells are hypothesized to balance replication speed with resource availability, but the factors and mechanisms regulating these divisions are not fully understood.
We want to understand how the environment and lifestyle of an organism shape the nuclear cycle control network. Do changes to the local environment alter the dynamics of nuclear cycles? Which mechanisms control nuclear multiplication in response to environmental cues? Is this regulation conserved between two organisms with different ecological niches?
To address these questions, we will take a comparative approach using two evolutionarily distant unicellular eukaryotes, the free-living slime mold Physarum polycephalum and the malaria parasite Plasmodium falciparum, which both have multinucleated life stages. We will compare the plasticity of nuclear cycling in these organism by using a combination of spatial transcriptomics, high-resolution imaging, basic cell biology techniques, and molecular genetics.
This project is supported by the Health + Life Science Alliance Heidelberg Mannheim awarded to Marie Jacobovitz, and is being carried out in collaboration with Markus Ganter at Heidelberg University Hospital.
Influence of environment on nuclear cycles


Evolution at the interface between chromosomes and the cell division machinery

During mitosis, the cell division machinery must rapidly and reproducibly partition the cell’s chromosomes through a stable but dynamic interaction between the centromere DNA, kinetochore complex, and spindle microtubules. Even though many components of the cell division machinery are highly conserved, eukaryotic chromosomes show striking diversity across species. Chromosomal elements such as centromeres range from genetically defined loci of fewer than 100 nucleotides in certain budding yeasts, to epigenetically defined regions of several megabases in metazoans. Additionally, the number of chromosomes (karyotype) can be remarkably different between species, even on extremely short evolutionary timescales. Together, these differences determine both the type and number of chromosomal regions that need to interact with proteins of the cell division machinery to ensure that each cell receives the correct number of chromosomes during mitosis or meiosis.
By combining experimental evolution (led by the Sherlock Lab at Stanford) and cell biological profiling (led by the Dey Group at EMBL), we are exploring how evolution acts on the interface between chromosomes and the cell division machinery. This project is funded by an EMBL/Stanford Bridging Excellence Fellowship awarded to Jana Helsen.
Coordination of microtubule organizing centers in space and time


Microtubule organizing centers (MTOCs) are key eukaryotic structures for microtubule nucleation and organization and can broadly be categorized into those with and those without a centriole. Centrioles fulfill two critical functions. In mitosis, they localize to the mitotic spindle pole aiding its organization. After cell division, centrioles can be repurposed as basal bodies at the plasma membrane where they are essential for flagella or cilia formation.
The function of centrioles/basal bodies (CBBs) correlates with subcellular localization, but is the spatiotemporal control required for this organelle to fulfill two distinct functions? How did the ancestral CBB evolve these functions? And why are different MTOCs not present simultaneously in a cell?
To address these questions, we will analyze and compare CBB repurposing strategies in two evolutionary distinct eukaryotes, the green alga Chlamydomonas reinhardtii and the slime mold Physarum polycephalum using state-of-the-art microscopy techniques. By interfering with the spatial and temporal coordination of the CBB we will try to elucidate the role of the subcellular location of this bifunctional organelle. Further, we will study how cells cope with several different MTOCs (centriolar and non-centriolar) at a time.
This project is supported by a Marie Sklodowska-Curie postdoctoral fellowship awarded to Caroline Simon and is being carried out in collaboration with the group of Niccolò Banterle at EMBL Heidelberg.
Impact of coenocytic lifestyle on the choice of mitotic strategies

Eukaryotes have evolved a spectrum of mitotic strategies, from open mitosis in animals which break down the nuclear envelope (NE) to closed mitosis in fungi that remodel the nucleus into a dumbbell before partitioning it. With most of the information gathered from a handful of model organisms, the factors driving this incredible diversity in nuclear reorganisation remain largely unknown.
We are interested in whether a coenocytic lifestyle, where many nuclei share a common cytoplasm, favours a more closed mitotic strategy. Does keeping the nuclear envelope intact allow many nuclei to exist and divide simultaneously in the same cell? Is there some interplay between nuclear envelope integrity and the nature of the microtubule organising centre (MTOC) and spindle architecture?
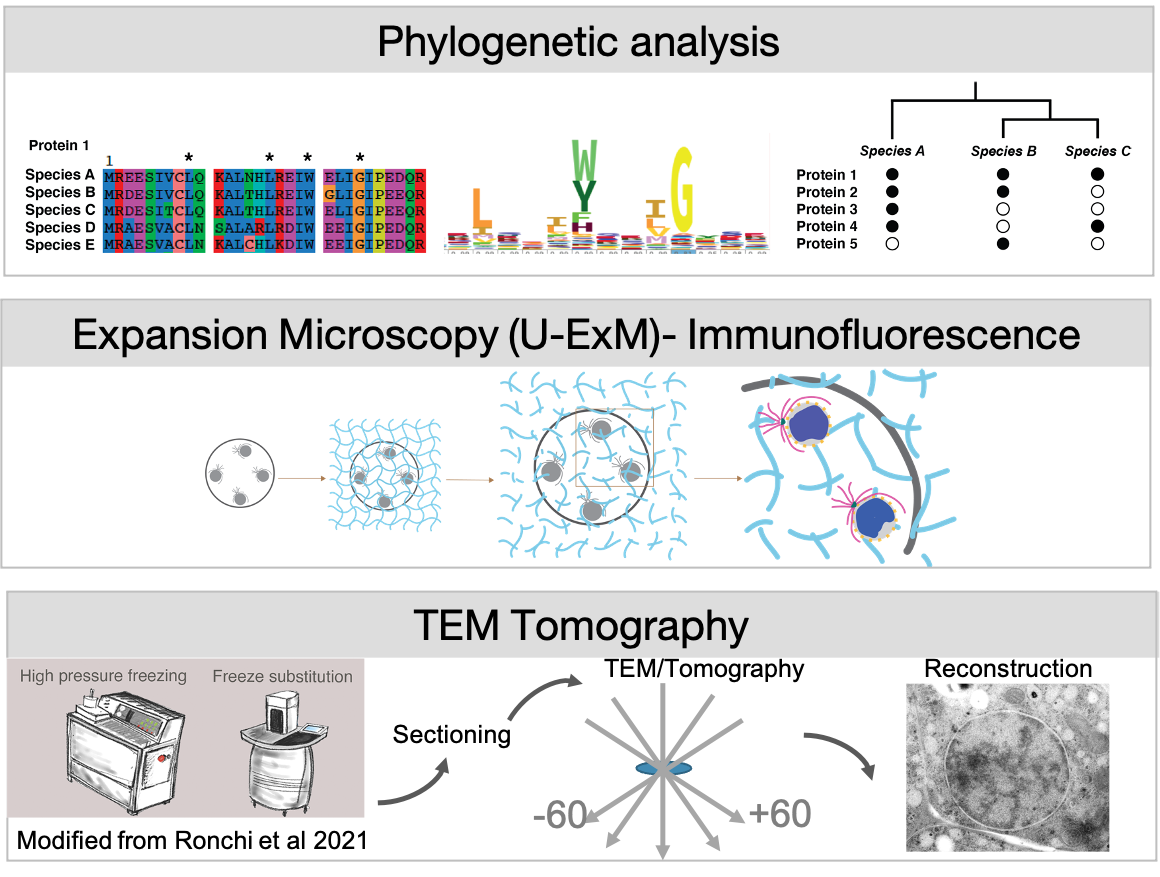
We address these questions in Ichthyosporea, close relatives of animals that exhibit a coenocytic lifestyle. Their unique position on the eukaryotic tree between our favourite animal and fungal models allows us to identify shared mechanisms of nuclear reorganisation. Using phylogenetics, and expansion microscopy and TEM-tomography we compare mitotic strategies between different Ichthyosporean species.
This project is supported by an EIPOD4 postdoctoral fellowship awarded to Hiral Shah, and is being carried out in collaboration with Yannick Schwab at EMBL and Omaya Dudin at EPFL, Lausanne.
TREC and Plan-ExM

We are joining the EMBL TREC (Traversing European Coastlines) expedition in 2023 and 2024!
We aim to create an atlas of planktonic ultrastructural diversity using Expansion Microscopy (ExM). By coupling expansion microscopy (Cryo-ExM and U-ExM) to 18s rRNA FISH we aim to identify species and image them at unprecedented 3D spatial resolution, with a long-term view to assess the repercussions of climate change on free-living microplankton populations.
To achieve this, we are pooling the expertise of several labs within (Schwab team, Saka Group) and outside of EMBL (Dudin Lab, Centriole Lab) .
Do you want to collaborate with us to use ExM on your favorite species? Contact us through this form


Diatom cell cycle modulation underlying bloom formation

Diatoms are single-celled organisms which are found ubiquitously throughout marine environments. Often present in large blooms, these cells collectively play an important role in the global biological pump. However, we have not deciphered the mechanisms underlying population dynamics of diatoms during bloom formation. This requires a closer look at individual cells within these populations.
To better understand how bloom formation plays out in terms of cell cycle modulation at the single-cell level, we are applying a multi-scale approach, observing cell size and morphology with cell cycle progression from one cell to its daughter cells, then the impact on population growth. We are working with the chain-forming species Chaetoceros decipiens, a species commonly found in blooms. High resolution, time-lapse observations of cells propagating in time and space along the chain allow us to evaluate how division rates change with respect to cell size and morphology. This work allows us to link single cell behavior and its emergent dynamics to gain a better understanding of how diatoms create such ecologically successful blooms.
This project is supported by an EIPOD-LinC Fellowship and is being carried out in collaboration with the Vincent lab and the Erzberger lab at EMBL.

Nucleolar dynamics during division and stress in fission yeasts
Besides its role in ribosome production, the nucleolus is involved in many other cellular functions, including mitosis regulation, mRNA transport, and stress response. It is a highly dynamic organelle with constant protein exchange with the rest of the nucleus and the cytoplasm. Changes in nucleolar behavior reflect the cell cycle stage and environmental conditions, and can also influence how the cell responds to them.
In our projects, we study how the nucleolus changes under stress and during cell division in two fission yeast species, Schizosaccharomyces pombe and Schizosaccharomyces japonicus. These species differ in nucleolar structure and in how they respond to stress, offering a good model for comparison. We combine live-cell imaging, genetics, and biochemical methods to follow nucleolar changes in normal and stressed cells.
We also explore how nucleolar division is controlled. In yeast, the nucleolus stays intact during mitosis, unlike in animal cells. This raises questions about how it splits and what proteins are involved. We use phylogenetic analysis to identify conserved factors, and examine how changes in protein networks affect nucleolar behavior.
Our goal is to build a detailed picture of nucleolar organization and its regulation, from cell cycle control to stress adaptation, and to understand how this varies across species.


From pixels to insights: Machine learning for ExM images

This is a collaboration with the Kreshuk lab
In our lab, we established working protocols for ultrastructure expansion microscopy (U-ExM) of eukaryotes across the tree of life. This technique allows us to physically expand biological samples, enabling faster and higher-resolution imaging while preserving the specificity of labeling.
However, the vast amount and complexity of data generated by U-ExM pose a major challenge: manual analysis is neither feasible nor scalable. To address this, we focus on developing advanced tools for data processing and analysis.
The project presented here establishes a machine learning framework to extract biologically meaningful insights from U-ExM images. By combining a pan-labeling strategy, which provides ultrastructure-level information, with unsupervised learning techniques that eliminate the need for manual labeling, we aim to make the analysis more efficient. Framed as a representation learning problem, this approach tackles a variety of tasks, including species classification, cell cycle and state analysis, phenotyping, morphometrics, and data integration
Mitotic transitions in the life cycle of Physarum polycephalum

During mitosis, nuclear envelope dynamics vary widely among organisms, with strategies ranging from complete nuclear envelope breakdown (open mitosis) to the retention of the envelope (closed mitosis). Interestingly, even closely related species such as Chromosphaera perkinsii and Sphaeroforma arctica (Ichthyosporeans) display different types of mitosis, suggesting that evolutionary transitions between these strategies have been common.
Physarum polycephalum, a unique amoebozoan, exhibits either open or closed mitosis in different stages of its life cycle. This dual strategy raises fundamental questions - How is the transition between open and closed mitosis regulated? How are cellular components, such as the nuclear envelope, MTOC, spindle fibers, and kinetochore, remodeled during these transitions? What are the transcriptomic and proteomic changes underlying this process? More broadly, how do these mechanisms inform our understanding of the evolutionary diversity of mitotic strategies?
To investigate this, we use state-of-the-art imaging techniques, including expansion microscopy, electron microscopy, and live-cell imaging coupled with transcriptomic and proteomic analyses.
